

Autor: Ing. Šimon Kertys, PhD.
Článok publikovaný v periodiku: Tichodroma 30: 3–9 (2018) ISSN 1337-026X
Úvod
Akustická identifikácia jedincov umožňuje ich sledovanie v priestore a čase, čo môže priniesť užitočné informácie o ich živote (Terry et al., 2005). U mnohých vtáčích druhov bolo zistené, že rôzne akustické vlastnosti môžu byť použiteľné pri identifikácii a následnom monitorovaní konkrétnych jedincov (napr. Darden et al., 2003; Terry et al., 2005), najmä u druhov so skrytým spôsobom života (napr. lelek lesný, Caprimulgus europaeus, Rebbeck et al. 2001; výr skalný, Bubo bubo, Grava et al. 2008).Táto neinvazívna metóda nevyžaduje odchyt, vizuálne pozorovanie a ani fyzické priblíženie k sledovaným jedincom. Nahrávanie spevu je všetko, čo je potrebné na identifikáciu (Budka et al., 2015). V prípade jariabka hôrneho sa jeho vokálnej individualite nevenovalo veľa autorov (Bergmann et al., 1975; Mulhauser & Zimmermann 2003). Doposiaľ získané výsledky z týchto prác poukazujú na možnosť identifikácie jedincov za pomoci štrukturálnych vlastností spevu tohto druhu. Cieľom tejto práce bol monitoring populácie jariabka hôrneho pomocou jednotlivých charakteristík ich spevu, ktoré boli použité na sledovanie jedincov tohto druhu v priestore a čase.
Materiál a metodika
Mapovanie jariabka hôrneho bolo realizované v severnej časti CHKO Horná Orava v katastrálnom území Rabčice a Oravská Polhora (Oravské Beskydy; 49°32’48.94″ s. š., 19°30’59.23″ v.d.; 750–1500 m n. m.; plocha cca 10 km2). Územie je súčasťou CHVÚ Horná Orava – SKCHVU008. Zbieranie údajov prebiehalo počas rokov 2015 – 2017. Celkovo bolo realizovaných za celé mapované obdobie 74 návštev.Vábenie prebiehalo vždy v jarných (marec, apríl, máj) a jesenných mesiacoch (september, október, november) počas toku. K vábeniu boli uprednostňované dni v mesiaci, kedy prevažovalo jasné a bezveterné počasie od svitu do 11 hod. a popoludní od 14 hod. do súmraku, kedy sme dosahovali najlepšie výsledky. Jariabky boli vábené pomocou vábničky imitáciou hlasu kohúta a sliepky, ktorou boli vyprovokované k hlasovým prejavom. Vábenie prebiehalo na vopred vytipovanom mieste a opakovalo sa v intervale každých 60 sekúnd päť krát za sebou, potom nasledovala dvojminútová odmlka a sledovanie okolia. Tento postup sa opakoval pravidelne v priebehu 15 – 20 minút. Ak sa v tomto časovom horizonte žiadny jedinec neozval, ani nepriletel, bolo potrebné sa potichu presunúť na ďalší vytipovaný bod, ktorý bol vzdialený max. do 300m od miesta vábenia. Počuteľnosť vábničky je závislá na viacerých faktoroch (vietor, typ porastu, typ vábničky), ale priemerná počuteľnosť vábenia je cca 150m. Po vyprovokovaní jedinca boli jednotlivé spevy nahrávané na diktafón typu Olympus DM-650, ktorý zaznamenával aj dátum a čas vábenia. Diktafóny neboli trvalo inštalované v teritóriách, ale spevy jednotlivých kohútov boli nahrávané priamo z ruky. Vzdialenosť nahrávania bola vždy závislá od vzdialenosti vyprovokovaného kohúta, od miesta vábenia (0 – 150m) a povahy kohúta. Vzdialenosť od nahrávaného jedinca mala vplyv aj na výslednú kvalitu nahrávky a zároveň aj na kvalitu spektogramu. Dĺžka nahrávok bola závislá od dĺžky vokálnej aktivity vyprovokovaného jedinca (1 – 16 spevov). Získané nahrávky boli následne vizuálne porovnávané v programe Raven Lite verzia 2.0.0. (http://www.birds.cornell.edu/brp/raven/RavenVersions.html#RavenLite). Priemerne bolo na jedného kohúta porovnaných 13 spevov a akustický podpis bol kombinovaný s miestom nahrávania. Spev jednotlivých kohútov bol zložený z viacerých elementov, ktoré sú rozdelené do viacerých fráz (obr. 1). Pre identifikáciu bolo zásadné usporiadanie jednotlivých elementov a celkový počet elementov, ktoré spolu tvoria „identifikačný prvok“( Bergmann et al., 1975). Počas porovnávania sme mali nastavené konštantné parametre pri všetkých spevoch. Pre fonetický prepis spevu sme použili spomalené prehrávanie, čo nám umožnilo presne rozpísať a spočítať počet elementov tvoriaci spev a rozdeliť elementy do jednotlivých fráz. V priebehu roka boli zaznamenávané aj nepriame pozorovania t.j. trus, stopy, popoliská a perie, ktoré sme použili na spresnenie území výskytu jednotlivých kohútov.
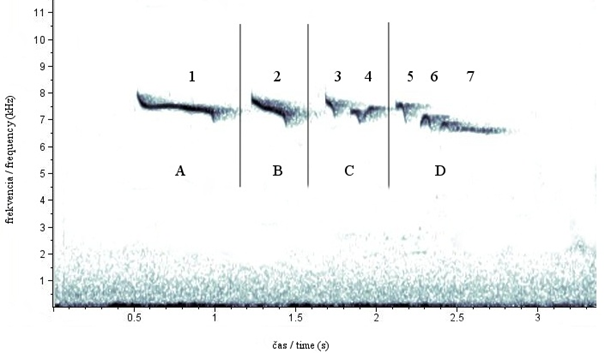
Obr. 1. Kompletný sonogram spevu jariabka hôrneho (Bonasa bonasia) – kohút K2 (CHKO Horná Orava, Slovensko). Písmená (A až D)zobrazujú frázy a čísla (1 až 7) jednotlivé elementy spevu.
Výsledky
Počas sledovaného obdobia bolo celkovo zaznamenaných a porovnaných 269 spevov zo 149 nahrávok. Z celkového počtu bolo na základe spevu identifikovaných 21 kohútov (obr. 2). V roku 2015 bolo porovnaných celkovo 64 spevov, z ktorých boli identifikované 4 kohúty na jar a 5 v jeseni, v 2016 roku z celkového počtu 129 spevov bolo identifikovaných 9 kohútov na jar a 3 v jeseni a v poslednom roku mapovania 2017 bolo porovnaných 76 spevov celkovo a identifikovaných 5 kohútov na jar a 7 v jeseni (tab.1). Spektrogramy spevu jednotlivých kohútov (obr. 2.) poukazujú na jednoznačné vizuálne rozdiely v ich zložení (usporiadanie a počet elementov).

Obr. 2. Spektrogramy poukazujúce na rozdielnu štruktúru spevu zaznamenaných kohútov počas mapovania.
Spev kohútov bol zložený zo 6 až 10 elementov (tab.1). Najviac kohútov malo spev zložený zo 7 elementov – 9 jedincov, 6 jedinci skladali spev z 8 elementov, v piatich prípadoch tvorilo spev 6 elementov a len v jednom prípade bol spev zložený z 10 elementov. Prvá fráza spevu (A fráza) kohútov nejavila výrazné individuálne znaky a bola tvorená jedným dlhým elementom. Výraznejšie individuálne znaky sa začali prejavovať až v ďalších častiach spevu (B, C a D frázy). B, C a D frázy sa vyznačovali výraznou stálosťou usporiadania elementov a umožnila identifikáciu kohútov. Na individuálne znaky nachádzajúce sa v týchto frázach poukazuje aj fonetický prepis spevu (tab. 1). Posledný obvykle dlhý element tvoriaci poslednú frázu ukončoval spev a menil jeho tonalitu. Celkový počet elementov bol v rámci jedincov stály a nemenil sa v priebehu sezón ani rokov.
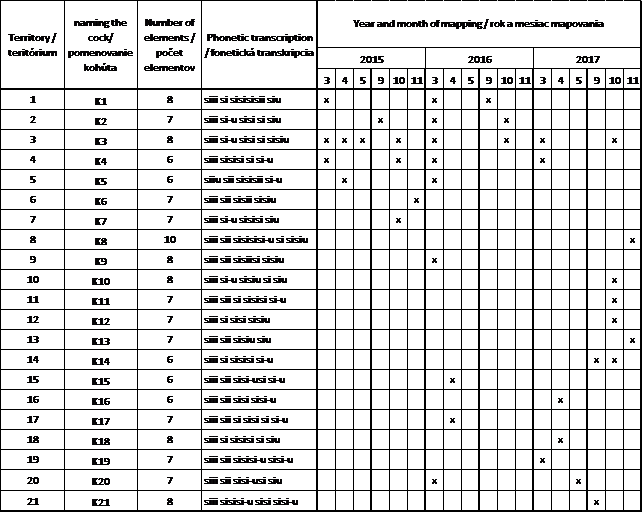
Tab. 1. Celkový prehľad zaznamenaných kohútov podľa rokov a mesiacov mapovania a charakteristiky ich spevu ( x – zaznamenanie spevu daného kohúta).
U7 kohútov bolo možné sledovať stálosť spevu (obr. 3.) ako aj presuny v rámci lokalít v priebehu sezón (jar a jeseň) a jednotlivých rokov. Kohúty z roka na rok a aj v priebehu sezón osídľovali rovnaké lokality a nemenili svoj spev. Na tento fakt poukazuje aj kohút K3, ktorý bol zaznamenaný v každej sezóne a roku mapovania (2015-jar a jeseň; 2016 jar a jeseň, 2017-jar a jeseň (tab. 1)) a v roku 2015 dokonca aj vo všetkých mesiacoch jarnej sezóny (marec, apríl, máj). V ďalších 6 prípadoch boli kohúty K1, K2, K4, K5, K14 a K20 zaznamenané minimálne v dvoch prípadoch v priebehu sezón, alebo rokov mapovania na rovnakých lokalitách. Aj tento fakt poukazuje na silnú stálosť a teritorialitu druhu. Maximálny posun kohúta v rámci svojej lokality bol zaznamenaný u samca K2, u ktorého bola vzdialenosť medzi záznamom spevu z 3/2015 a 3/2016 až 480m. V prípade ďalších kohútov, ktoré boli zaznamenané opakovane v priebehu mapovaného obdobia (K1, K3, K4, K5, K14 a K20) boli presuny (sezóne, medziročné) v rámci svojho teritória do 400m.
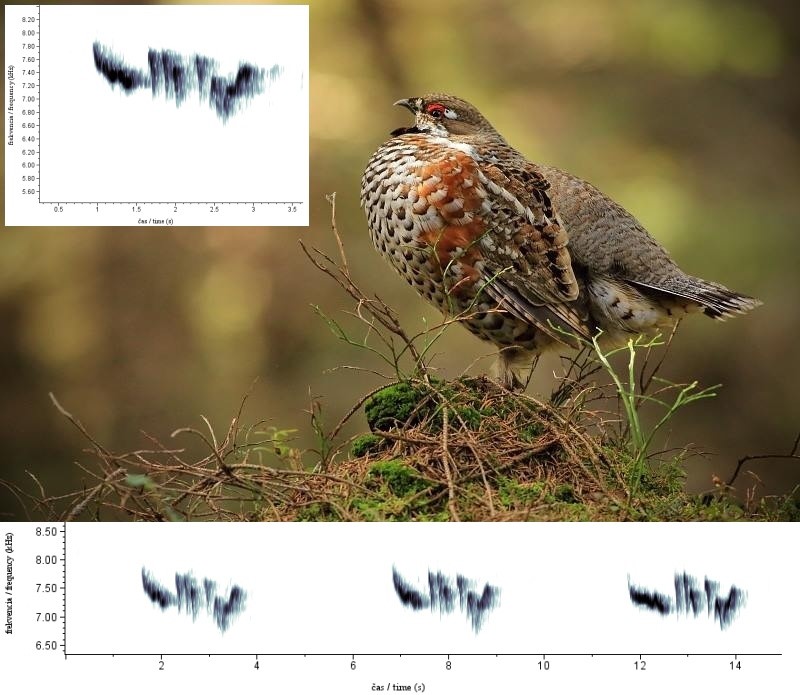
Obr. 3. Spektrogramy kohúta K4 z dňa 1.10.2015, ktoré poukazujú na stálosť spevu z jednej nahrávky. Dĺžka prestávky medzi spevmi bola v tomto prípade graficky skrátená. (foto: Šimon Kertys)
Pri bližšom vizuálnom porovnávaní spektrogramov jednotlivých kohútov možno zistiť, že počet a usporiadanie elementov tvoriacich spev je identické nielen v krátkom časovom horizonte (obr. 3), ale nemení sa ani v priebehu rokov (obr. 4).
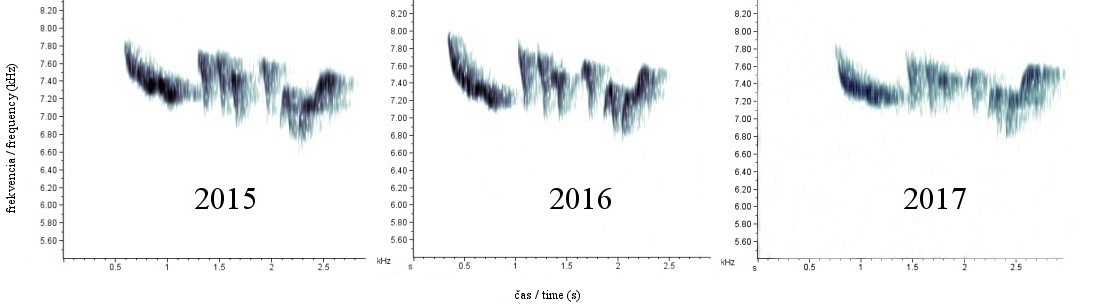
Obr. 4. Porovnanie spektrogramov kohúta K4 z rokov 2015 – 2017.
Diskusia
Výsledky práce poukazujú na možnosť identifikácie kohútov jariabka hôrneho v priestore a čase, na základe charakteristík ich spevu. Na tento fakt poukazuje aj práca Mulhauser & Zimmermann (2003), v ktorej autori identifikovali za pomoci nahrávok spevu 13 kohútov jariabka hôrneho. Spektrogramy spevov rôznych kohútov poukazovali na jednoznačné rozdiely medzi jedincami. Tiež zistili, že spev je väčšinou zložený zo 6 až 11 elementov, čo potvrdzuje aj táto práca, kde bol spev zložený zo 6 až 10 elementov. Podľa týchto autorov nie je ani na prvý pohľad malý počet elementov tvoriaci spev problémom pri tvorbe rozdielov v jednotlivých spevoch kohútov. Napríklad uvádzajú, že pre spev zložený zo 6 elementov má 16 možností variant, spev tvorený 7 elementami až 36 variant a pre spev tvorený z 8 elementov má až 64 variant. Bergmann et al. (1975) študovali variácie spevu v populácii jariabka hôrneho v Białowieżi NP (Poland) prostredníctvom sonografických metód. Celkovo zhodnotili 250 spevov z 15 rôznych jedincov. Zistili, že každý kohút v študovanej populácii, by mohol byť identifikovaný na základe individuálnych vlastností jeho spevu. Autori tiež poukázali, že variabilita spevu medzi jedincami tohto druhu sa dosiahne buď dodatočnými prvkami, nedostatkom prvku, odlomením alebo spájaním prvku. Swenson (1991) v svojej práci zisťoval hustotu teritórií jariabka hôrneho v jarnom a jesennom období za pomoci vábničky. Počet zaznamenaných samcov preukázateľne lineárne koreloval s počtom známych teritórií samcov na študovanom území. Zistil, že až 82 % teritoriálnych samcov odpovedalo na vábenie a bolo zaznamenaných. V našom prípade bolo z 26 predpokladaných teritórií potvrdených 21 v rámci sledovaného územia, z čoho celkovo zareagovalo na vábenie a bolo zaznamenaných až 80% kohútov. Jednotlivé teritória sme odhadli na základe zozbieraných pobytových znakov, ktoré sme zaznamenávali celoročne (stopy, trus, nocoviská, popoliská, perie atď.). Na možnosť využitia akustických vlastností spevu pre identifikáciu poukazuje viacero autorov (Grava et al. 2008, Klenova et al., 2008, Budka et. al., 2015, Petrusková et al. 2016) na rôznych druhoch. Nie je však možné úplne porovnávať jednotlivé druhy, keďže rozdiely v speve môžu byť výrazné od komplexných a bohatých spevov až po jednoduchú vokalizáciu, ako je to v našom prípade. Aj napriek tomu v prípade vokálnej identifikácie je potrebné vždy zistiť jednotlivé rozdiely v speve a posúdiť stabilitu zistených rozdielov z roka na rok. Aj v našej práci bolo možné porovnať v 7 prípadoch stabilitu usporiadania a počtu jednotlivých elementov spevu a na základe toho identifikovať jedincov v priestore a čase. Výsledky práce pomohli identifikovať individuálne znaky spevu, ktoré poslúžili ako individuálny akustický podpis. Za pomoci individuálneho akustického podpisu dokážu niektoré druhy rozoznávať známeho a neznámeho samca na základe jediného spevu (Lambrechts & Dhondt 1995) a niektoré na základe jediného spevu rozlišujú aj známych samcov medzi sebou (napr. strnádlik spevavý, Melospiza melodia, Stoddard et al.1990; kolibkárik čipčavý, Phylloscopus collybita, Jaška et al. 2015). Jednotlivé druhy môžu využívať ako individuálny akustický podpis celý spev, ktorý sa svojimi vlastnosťami odlišuje od spevu ostatných samcov (napr. pamuchár jelšový, Empidonax alnorum, Lovell& Lein 2005),môžu identitu určiť aj na základe vyčlenenia jednej časti spevu (napr. strnádlik bielohrdlý, Zonotrichia albicollis, Brooks&Falls 1975; strnádlik Pugetov, Zonotrichia leucophrys pugetensis, Nelson &Poesel 2007) a v niektorých príkladoch je ukrytá identita na vyššej úrovni, ako napr. usporiadanie slabík v speve (napr. škovránok poľný, Alauda arvensis, Briefer et al. 2008; ľabtuška lesná, Anthus trivialis , Petruskováet al. 2016). Táto jednoduchá metóda, môže byť nápomocná pri zisťovaní veľkosti populácie, priestorových nárokov, miery prežitia ako aj dlhovekosti jariabka hôrneho. Rozdiely v komponovaní spevu medzi jednotlivými kohútmi, boli výrazné a sú veľmi dobre viditeľné. Veľmi nápomocné je aj silná stálosť a teritorialita druhu čo nám dáva veľkú pravdepodobnosť, že jedinec bude obhajovať rovnaké územie a je možné ho tam zaznamenať v priebehu viacerých rokov.
Použitá literatúra
Bergmann H. H., Klaus S., Müller F. & Wiesner J. 1975: Individualitätund Artspezifität in den Gesangsstropheneiner Population des Haselhuhns (Bonasabonasia L., Tetraoninae,Phasianidae). — Behaviour 55: 94–114.
Briefer E., Aubin T., Lehongre K. & Rybak F. 2008: How to identify dear enemies: the group signature in the complex song of the Skylark Alauda arvensis. — Journal of Experimental Biology 211: 317–326.
Brooks R. J. & Falls J. B. 1975: Individual recognition by song in White-throated Sparrows. III. Song feature used in individual recognition. — Canadian Journal of Zoology 53: 1749–1761.
Budka M., Wojas L. & Osiejuk T. S. 2015: Is it possible to acoustically identify individuals within a population? — Journal of Ornithology 156: 481 – 488.
Darden S. K., Dabelsteen T. & Pedersen S. B. 2003: A potential tool for swift fox (Vulpes velox) conservation: individuality of long-range barking sequences. — Journal of Mammalogy 84: 1417–1427.
Grava T., Mathevon N., Place E. & Balluet P. 2008: Individual acoustic monitoring of the European Eagle Owl Bubo bubo. — Ibis 150: 279–287.
Jaška P., Linhart P. & Fuchs R. 2015: Neighbour recognition in two sister songbird species with a simple and complex repertoires – a playback study. — Journal of Avian Biology 46: 151–158.
Klenova A. V., Volodina I. A. & Volodina E. V. 2008: Duet structure provides information about pair identity in the red-crowned crane (Grus japonensis). — Journal of Ethology 26: 317–325.
Lambrechts M. M. & Dhondt A. A. 1995: Individual voice discrimination in birds. — In: Power D. M. (ed.) 1995: Current Ornithology. Vol. 12 Plenum Press, New York: 115–139.
Lovell S. F. & Lein M. R. 2005: Individual recognition of neighbours by song in a suboscine bird, the Alder Flycatcher Empidonax alnorum. — Behavioral Ecology and Sociobiology 57: 623–630.
Mulhauser B. & Zimmermann J. L. 2003: Recognition of male hazel grouse Bonasa bonasia by their song. — Bulletin de la Société Neuchâteloise des SciencesNaturelles126 (2): 107–119.
Nelson D. A. & Poesel A. 2007: Segregation of information in acomplex acoustic signal: individual and dialect identity in White-crowned Sparrow song. —Animal Behaviour 74: 1073–1084.
Petrusková T., Pišvejcová I., Kinštová A., Brinke T. & Petrusek A. 2016: Repertoire-based individual acoustic monitoring of a migratory passerine with complex song as an efficient tool for tracking territorial dynamics and annual returns rate. — Methods in Ecology and Evolution 7: 274–284.
Rebbeck M., Corrick R., Eaglestone B., Stainton C. 2001: Recognition of individual European Nigtjars Caprimulgus europaeus from their song. Ibis 143: 468–475.
Stoddard P. K., Beecher M. D., Horning C. L. & Willis M. S. 1990: Strong neighbour stranger discrimination in Song Sparrows. — Condor 92: 1051–1056.
Swenson J. E. 1991: Evaluation of a density index for territorial male hazel grouse Bonasa bonasia in spring and autumn. — Ornis Fennica 68:57–65.
Terry A. M. R., Peake T. M. & McGregor P. K. 2005: The role of vocal individuality in conservation. — Frontiers in Zoology 2: 10.